Why Johnny can’t read (but Jane can)
Reading is not a skill that comes naturally. Unlike learning spoken language, which the human brain has evolved to absorb almost effortlessly, learning to read is a protracted and difficult process. It involves the categorical association of arbitrary visual symbols with phonemes and also the ability to break words down into component phonemes. It thus relies on an integration between visual and auditory processes, combining spatial and temporal information, within a learned linguistic context. The fact that reading is such a specialized and integrative skill may partly explain why it can be selectively impaired in people of otherwise normal intellectual abilities.
Dyslexia, as this type of selective reading difficulty is called, is quite common, affecting anywhere from 5% to 20% of children, depending on the criteria used in its diagnosis. Cohort studies which directly tested all individuals have found that dyslexia is about twice as common in boys as in girls. This is not unusual; many psychiatric disorders show a different rate of occurrence between the sexes, with autism, schizophrenia and attention-deficit hyperactivity disorder all being more common in males, while depression and some other non-psychiatric conditions, such as synaesthesia, are more common in females. The study of dyslexia is providing some insights into how the interaction between predisposing factors and sex can influence phenotypic outcome.
First, some clues as to what the underlying cause may be. Dyslexia is highly heritable and in recent years a number of genes have been linked to it, through unbiased genetic mapping approaches. Remarkably, all of these genes are involved in some way in controlling cell migration in the cerebral cortex. This astonishing convergence parallels neuroimaging and neuropathology studies which have found a high rate of subtle malformations of the cerebral cortex in brains of dyslexic subjects. These include small clusters of misplaced neurons (termed nodular heterotopia), as well as small infoldings of the cortical sheet (microgyria). Disruption in the normally highly regular organization of the cortex may thus underlie this disorder.
One hypothesis to explain how this occurs is that these heterotopia and microgyri disrupt communication between cortical areas in a way that impairs the integrative processes required for reading.
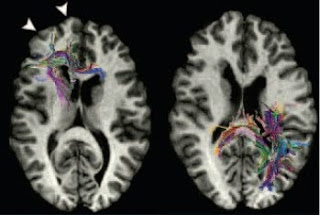
A study from the lab of Christopher Walsh provides direct evidence supporting this idea. He and his colleagues studied a rare, inherited condition, called periventricular nodular heterotopia, which is characterised by the presence of misplaced clusters of neurons within the white matter near the cerebral ventricles, and which is also associated with very high rates of dyslexia. Using diffusion tensor imaging to follow specific axonal tracts within the brain, the authors found that these heterotopia did indeed interrupt the structural connections that passed through the affected regions (while tracts in parts of the brain without heterotopia were normal).
But the impact of such displaced neurons is not limited to the cortex itself. A fascinating set of studies from Albert Galaburda and colleagues with an animal model of microgyria has revealed secondary consequences on other parts of the brain that connect to these regions, in particular the thalamus. In young rats, very localized microgyria can be induced by application of a freezing probe to the surface of the brain (in this case to the parietal cortex). These lesions resulted in defects in auditory processing, specifically of very fast temporal sequences, which may parallel some of the auditory processing defects seen in dyslexia, especially in phonemic processing. The lesions also resulted in secondary changes in the medial geniculate nucleus, the auditory centre of the thalamus which connects to the parietal cortex. Most remarkable, however, was that both the histological changes in the thalamus and the defects in auditory processing were much more pronounced in male rats than in females. When females were treated with testosterone and lesioned they showed similar secondary effects to male rats. It is known that testosterone regulates proliferation, cell death and connectivity during development, but it is also involved in various processes of neuroplasticity, which may explain the differences in secondary effects observed here.
The sex difference in this case thus involves not the primary defect itself but the way in which the brain responds to it. As many psychiatric disorders involve an interaction between primary neurodevelopmental insults and ongoing neuroplasticity processes, this mechanism may be of much wider relevance in explaining the different susceptibility of male and female brains to various conditions.
Chang, B., Katzir, T., Liu, T., Corriveau, K., Barzillai, M., Apse, K., Bodell, A., Hackney, D., Alsop, D., Wong, S., & Walsh, C. (2007). A structural basis for reading fluency: White matter defects in a genetic brain malformation Neurology, 69 (23), 2146-2154 DOI: 10.1212/01.wnl.0000286365.41070.54
Rosen GD, Herman AE, & Galaburda AM (1999). Sex differences in the effects of early neocortical injury on neuronal size distribution of the medial geniculate nucleus in the rat are mediated by perinatal gonadal steroids. Cerebral cortex (New York, N.Y. : 1991), 9 (1), 27-34 PMID: 10022493



{kind=link}
Fascinating. It will be interesting to see how this plays out in future research.
ReplyDeleteI think Christopher Walsh gave a lecture in my neuroscience class.
Thanks Livia, for your comment. One of the interesting questions for the future will be whether this kind of cell migration defect underlies all cases of dyslexia or only a subset and whether such a subset might have some distinctive clinical features. There are currently debates about whether such subtypes exist, defined by various characteristics - defects in phonemic processing, defects in fine motor control (including eye-scanning), cerebellar defects, and others.
ReplyDeleteHello????????????????????(me scratching my head)?
ReplyDeleteSo if a person is brain damaged later in life is it the same thing that would cause their dyslexia?
I,m not trying to sound ignorant here but rather from a view point of a person trying to understand their mind and its weird ways instead of becoming a neurologist.
I can re-read these words and "feel" quite a bit of what is said.
My neurons misfire fairly often and only recently (2yrs) have they caused problems like dyslexia .
I over come a lot of it by using a trick that I taught a former friend is to read aloud to your self if necessary and my response time in comprehending something speeds up considerably.
I understand the probe doing minor damage on a brain then seeing the results but what is the difference when a brain has the same problems (auditory processing) but it olny temporary? Rick
seems impossible,almost five years since my father was diagnosed with shoulder problems,and this with a strong treatment,but now is having a hard hit because when dysfunction was a problem in the past but returned and want to take generic viagra and do not know if this may affect your treatment and if in the same manner and shall take effect not do?
ReplyDeleteI had a friend to have this condition, the doctor said that it happen because he used to Buy Generic Viagra a lot and eat a lot of fat food and stuff.
ReplyDeleteGreat blog Kevin...I'm a regular visitor. It was my understanding that dyslexia doesn't discriminate and that it affects both sexes equally. The view that boys suffer more stemming from the idea that boys are more likely to demonstrate frustrated/disruptive behaviour etc. more so than girls who will tend to internalise the problem and therefore delay diagnosis?
ReplyDeleteThere is definitely so much research that needs to be done with this so much here. So many people that deal with this so much and need the help with it. I hope they will be able to tackle this. Mortgage brokers Toronto
ReplyDeleteReally you deserve a round of applause for this highly knowledgeable post in this segment of medical transcription toronto. Thank you for such an informative post.
ReplyDelete